【大国粮策】转座子是植物适应性进化的幕后推手吗?
在植物基因组中,除了那些我们熟悉的控制开花时间、抗病能力和产量的“常规基因”,还有一类特别的“跳跃基因”——转座子。它们就像基因组里的“唱跳练习生”,随时可能换个位置,闪亮登场。转座子有时悄悄“插队”到某个基因附近,改变基因的表达节奏;有时又“搬家”到新的位置,带来全新的表观遗传变化。科学家曾经认为它们只是无关紧要的“杂音”,但事实证明,它们却频频在植物进化舞台上“C 位出道”。
从“疯狂”到诺贝尔奖——转座子是如何被发现的?
转座子的“唱跳”天赋并不是最近才被发现的。早在1951年,美国女科学家芭芭拉·麦克林托克(Barbara McClintock)就在冷泉港研讨会上首次提出了基因“跳跃”的理论。当时,她在研究玉米籽粒颜色的遗传规律时,发现某些基因的位置居然会在不同世代之间“变来变去”,就像会自己挪动的拼图块。她注意到,这种跳动与籽粒上出现的斑点花纹有直接关系——有的籽粒颜色深浅不一,就是因为转座子插入或离开了与色素合成相关的基因。
这一发现颠覆了当时“基因位置固定不变”的主流观点,但在那个年代,她的理论并未立刻被广泛接受,甚至有人觉得她的想法“太疯狂”。直到数十年后,分子生物学的进步才让科学界证实了她的预言。1993 年,麦克林托克因“发现可移动的遗传元件”获得诺贝尔生理学或医学奖,成为这一领域最闪亮的“星探”和伯乐。
“减益”与“加成”——转座子的双面特性
转座子的“跳跃”有时像给植物加了debuff(敏感性增加),可能削弱植物在某些环境中的竞争力;有时则像触发了 buff(抗寒、抗旱),帮助它们在恶劣环境中稳定生长。
具体来说,前人研究中发现,番茄slDCL2a基因中一个罕见的MuDR转座子插入,导致slDCL2a基因表达显著下降,使携带MuDR插入的等位基因的植物更易感染RNA病毒,从而使其更易感染晚疫病。另外,番茄中还发现一个COPIA转座子插入Ph-3基因的外显子区域,导致Ph-3基因(抗疫霉菌)转录本截短、表达量降低,这种变化可能增加植物对疫霉菌的敏感性。
除了负面影响,转座子也能带来积极的变化。近期研究人员通过全基因组图谱构建和重测序数据分析,在模式植物水稻中发现了超过 3 万个转座子插入多态性(TIP)位点,并利用转座子插入多态性全基因组关联分析方法锁定 OsCACT 等新的耐冷候选基因。功能验证结果表明,这些基因显著提升了水稻在低温环境下的存活率。这项成果不仅为水稻培育提供了新的靶点,也凸显了转座子在适应性进化中作为“幕后推手”的角色——它们犹如为植物悄悄设计的一件“防寒外套”。
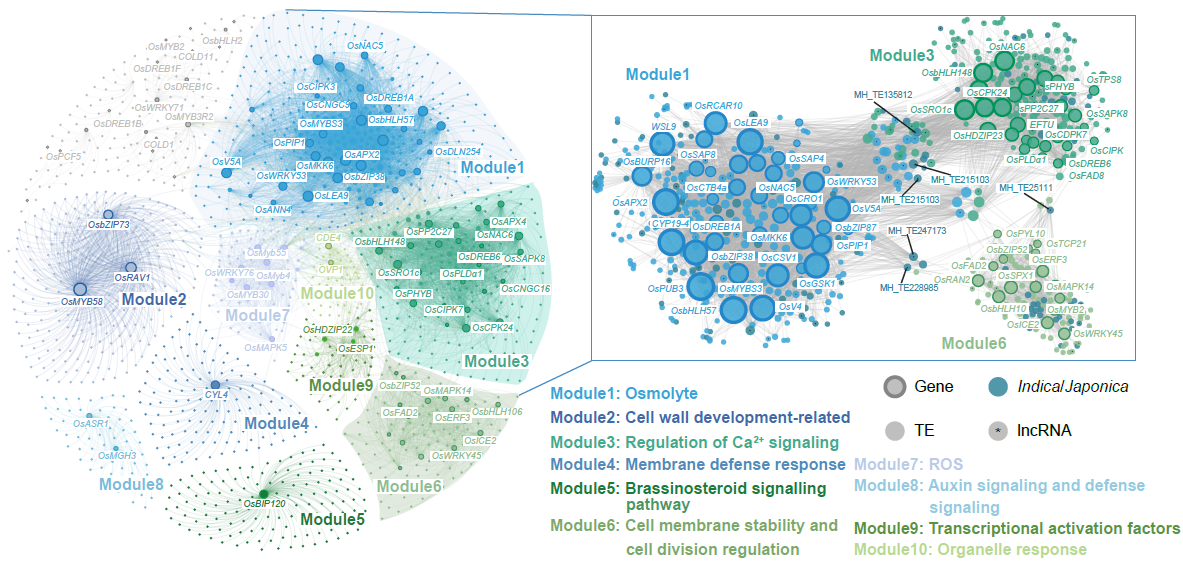
正是这种“正负双刃剑”的特性,使得转座子成为推动植物适应性进化的重要变量——它们既可能带来突破性的生存优势,也可能制造不利因素,从而在自然选择的筛选下,塑造出更适应环境的植物群体。
适应性进化:从“随机跳跃”到“生存智慧”
转座子的跳跃本质上是一种随机变化,但在长期环境筛选下,有利的跳跃会被保留下来,不利的则被淘汰。比如,长末端重复序列(LTR)逆转座子插入BnSHP1.A9的启动子区域,通过DNA甲基化表观遗传沉默BnSHP1.A9的表达,从而增强了油菜的豆荚抗裂性。hAT、MITE和LINE转座子的插入还与油菜开花时间变化相关。例如,BnaA10.FLC基因第一个外显子中插入的LINE导致功能丧失,春型油菜(SOR)春化需求减弱或不需春化;MITE插入启动子区增强BnaA10.FLC表达,使冬型油菜(WOR)需较强春化;而hAT插入使半冬型油菜(SWOR)的春化需求介于两者之间。
转座子通过引入新的基因调控机制,改变基因表达的时空模式,赋予植物更强的环境适应能力。这些基因组中的“偶然改造”,就像植物的“生存手册”,历代积累,形成丰富的遗传智慧。植物不仅被动适应环境,更像拥有一套灵活的“基因工具箱”,能在环境变化时迅速调整自身生理状态。
因此,适应性进化不仅是随机突变的缓慢积累,更包含了转座子等能够“自我改造”的动态过程,是自然选择与基因组灵活性共同作用的结果。正是这份“生存智慧”,成就了植物的多样性与广泛适应性。
转座子与农业科技的结合
随着基因组学、转录组学、表观遗传学以及单细胞测序的发展,我们现在不仅能追踪转座子的历史“足迹”,还能实时观察它们在不同环境下的“活动轨迹”。更令人期待的是,科学家们正尝试将转座子“驯化”为可控的育种工具。
研究人员整理了多态性转座子信息数据库,供全球科研人员在线查询和利用,作为分子标记辅助选育更优质品种。未来,借助基因编辑等技术,或可在特定环境下精准激活这些转座子,使作物“快速升级”,更好应对干旱、高盐、病害等胁迫。
在全球气候变化的大背景下,转座子作为基因组中具有高变异性和调控潜力的重要因子,有望成为保障粮食安全的关键力量。我们不再被动等待自然选择的缓慢筛选,而是可以主动调控这些转座子,为现代农业生产“排兵布阵”,实现更加精准高效的作物改良。
参考文献:
[1] B. McClintock. The origin and behavior of mutable loci in maize, Proc. Natl. Acad. Sci. U.S.A. 36 , 344-355 (1950). https://doi.org/10.1073/pnas.36.6.344
[2] Domínguez, M., Dugas, E., Benchouaia, M. et al. The impact of transposable elements on tomato diversity. Nat Commun 11, 4058 (2020). https://doi.org/10.1038/s41467-020-17874-2
[3] YQ Qian, ZW Zhou, OY Tianmin, et al. Pangenome analysis of transposable element insertion polymorphisms reveals features underlying cold tolerance in rice. Nat Commun (2025).
[4] J Liu, RJ Zhou, WX Wang, et al. A copia-like retrotransposon insertion in the upstream region of the SHATTERPROOF1 gene, BnSHP1.A9, is associated with quantitative variation in pod shattering resistance in oilseed rape. J. Exp. Bot. 71, 5402–5413 (2020). https://doi.org/10.1093/jxb/eraa281
[5] Song, JM., Guan, Z., Hu, J. et al. Eight high-quality genomes reveal pan-genome architecture and ecotype differentiation of Brassica napus. Nat. Plants 6, 34–45 (2020). https://doi.org/10.1038/s41477-019-0577-7
[6] Z Liu, TZ Wang, L Wang, et al. RTRIP: a comprehensive profile of transposon insertion polymorphisms in rice. Plant Biotechnol. J. 18, 2379-2381 (2020). https://doi.org/10.1111/pbi.13425
策划:谢芸
作者:宋佳明 西南大学研究员、博士生导师
审核专家:陈玲玲 崖州湾国家实验室教授、博士生导师
图文简介